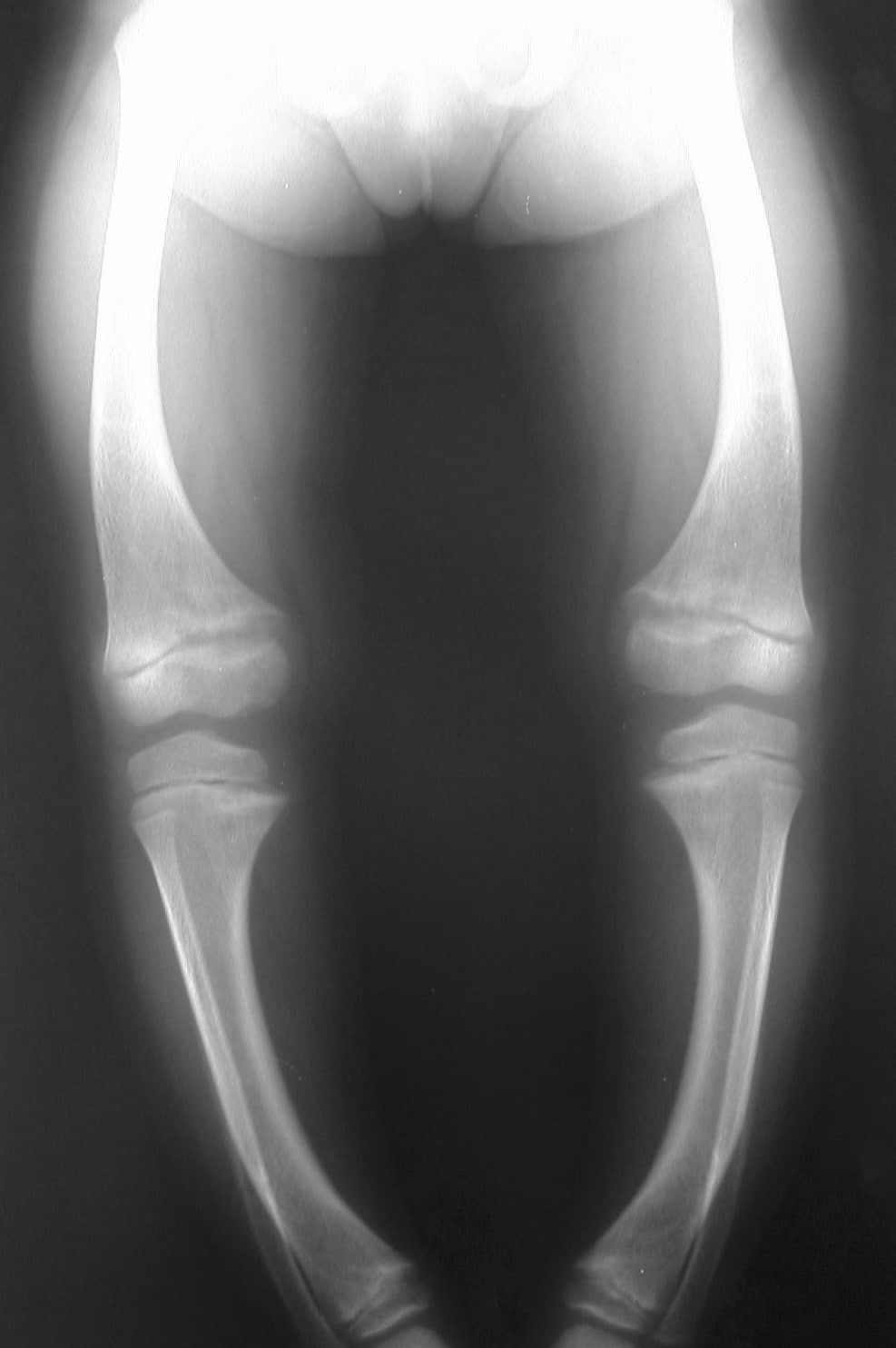
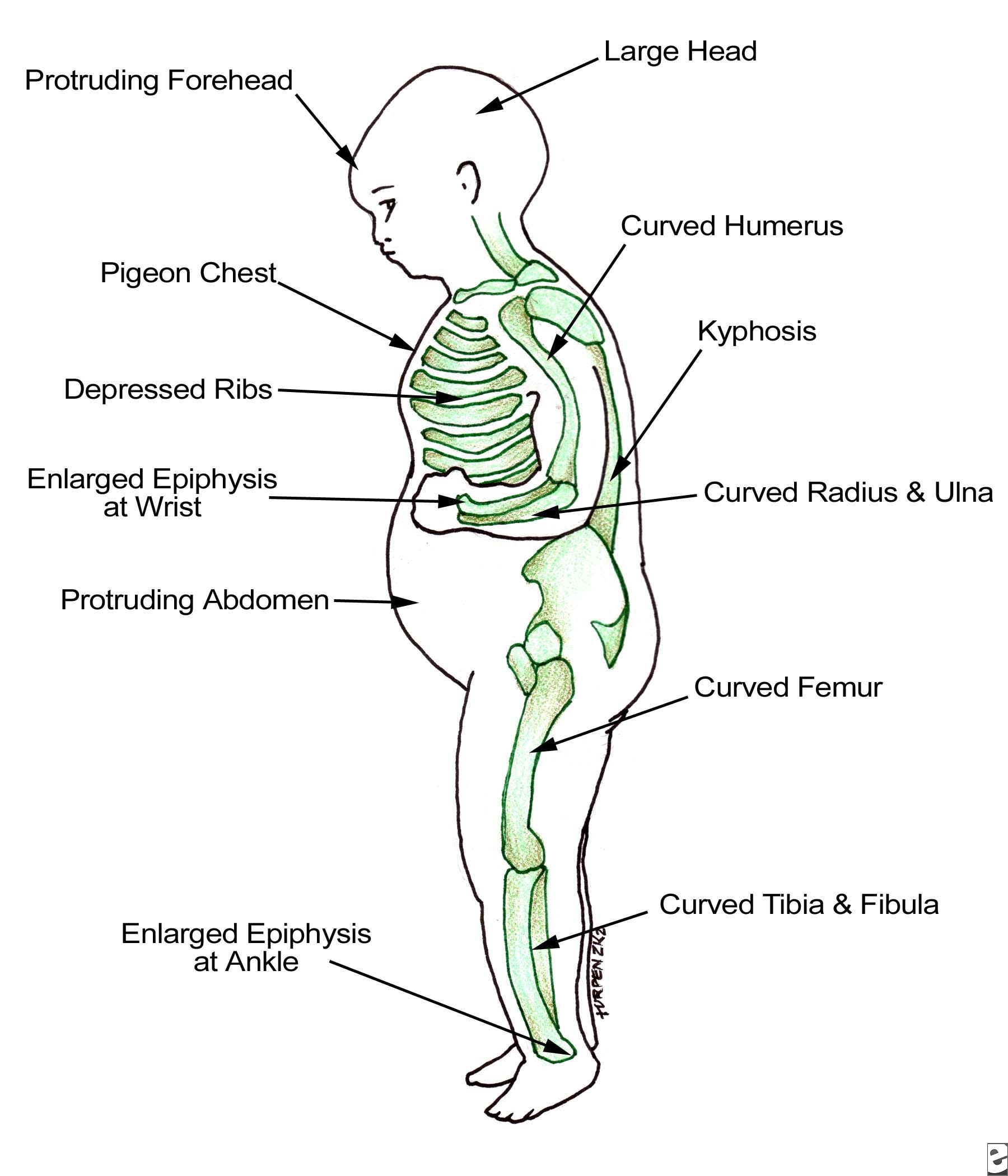
صد نکته کنکوری در بیولوژی مولکولی
1- آلكاپتونوريا يك نوع بيماري ارثي است ( مربوط به ژن و DNA )
2- ادرار افراد مبتلا به آلكاپتونوريا در مجاورت هوا سياه رنگ مي شود ( واكنش هموجنتيسيك اسيد با هوا )
3- در افراد مبتلا به آلكاپتونوريا آنزيم تجزيه كننده هموجنتيسيك اسيد وجود ندارد .( نظر آرچيبلد گرو )
4- اين بيماري رابطهي بين يك ژن و يك نقص آنزيمي را نشان مي دهد.
5- كپك نوروسپورا نوعي قارچ هاپلوئيد (يوكاريوت و n كروموزومي ) مي باشد ، كه در زمان كوتاهي تعداد زيادي هاگ توليد مي كند.
6- محيط كشت حداقل شامل انواع نمك ها – كمي شكر و يك نوع ويتامين به نام بيوتين است .
7- بيدل و تيتم از اشعه X براي ايجاد جهش در هاگ ها استفاده كردند .
8- بيدل و تيتم هاگ هايي كه در محيط حداقل نمي توانستند رشد كنند را جهش يافته ناميدند .
9- مسير متابوليكي پيشنهادي براي توليد آرژينين با توجه به آزمايش هاي بيدل و تيتم به صورت زير است .
X ارنيتين سيترولين آرژينين
10- جهش يافته هاي نيازمند آرژينين 3 دستهاند :
a. گروهي كه با اضافه كردن ارنيتين يا سيترولين و يا آرژينين به محيط كشت حداقل رشد مي كردند. در اين گروه آنزيم شمارهي 1 وجود ندارد.
b. گروهي كه با اضافه كردن سيترولين يا آرژينين به محيط كشت حداقل رشد مي كنند. در اين گروه آنزيم شماره ي 2 وجود ندارد.
c. گروهي كه فقط با اضافه كردن آرژينين به محيط كشت حداقل رشد مي كنند. اين گروه آنزيم شمارهي 3 را ندارند.
11- بيدل و تيتم نتيجه گرفتند كه هر ژن از طريق كنترل توليد يك نوع آنزيم اثر خود را اعمال مي كند.
12- به دليل اينكه 1) بسياري از ژن ها پروتئين هايي را رمز مي كنند كه آنزيم نيستند و 2) بسياري از پروتئين ها از چند زنجيرهي پلي پپتيدي تشكيل شدهاند كه توليد هر زنجيره را يك ژن خاص رهبري مي كند ، نظريه يك ژن – يك آنزيم به نظريه ي يك ژن يك رشتهي پلي پپتيدي تغيير كرده است.
13- منظور از رمز علائمي است كه براي ذخيره سازي و انتقال اطلاعات به كار مي رود.
14- اطلاعات در DNA به صورت رمز است ؛ زبان رمز DNA فقط داراي چهار حرف است ( 4 نوع نوكلئوتيد A , T, C , G )
15- رمز هاي DNA تعيين كننده ي نوع و ترتيب اسيد هاي آمينه در پروتئين ها مي باشند.
16- در پروتئين ها در كل 20 نوع اسيد آمينه وجود دارد.
17- رمزهاي DNA براي هر اسيد آمينه بايد حداقل سه حرفي باشند. ( توالي سه نوكلئوتيد)
18- با در نظر گرفتن سه حرفي بودن رمز ها در DNA ، در كل 64 نوع رمز در DNA براي 20 نوع اسيد آمينه وجود دارد.
19- به رمز هاي DNA كد و به رمز هاي DNA كدون و به ضد رمز هاي tRNA آنتي كدون گفته مي شود.
20- رابطه ي بين DNA و سيتوپلاسم را مولكولي به نام RNA برقرار مي كند.
21- دلايل وجود RNA عبارتند از 1) در يوكاريوت ها DNA در هسته است ولي پروتئين سازي در سيتوپلاسم ، پس نياز به يك واسطه وجود دارد. 2) سلول هايي كه فعاليت پروتئين سازي در آنها بيشتر است ، مقدار بيشتري هم RNA دارند. 3) RNA هم در هسته و هم در سيتوپلاسم يافت مي شود.
22- RNA يك مولكول تك رشته اي است كه در آن به جاي نوكلئوتيد T ، نوكلئوتيد U وجود دارد و قند نوكلئوتيدهاي آن ريبوز است.
23- در همه ي انواع سلول ها سه نوع RNA وجود دارد كه عبارتند از
a. mRNA كه اطلاعات DNA را به ريبوزوم ها در سيتوپلاسم حمل مي كند.(RNAي پيك يا پيامبر)
b. tRNA كه آمينو اسيد ها را به ريبوزوم منتقل مي كند ( RNAي ناقل )
c. rRNA كه همان RNAي ريبوزومي است و در ساختمان ريبوزوم شركت دارد .
24- ساخته شدن RNA از روي DNA را رونويسي گويند.
25- رونويسي اولين قدم براي ساختن پروتئين است.
26- رونويسي با كمك آنزيمي به نام RNA پلي مراز صورت مي گيرد.
27- سلول هاي پروكاريوتي فقط يك نوع RNA پلي مراز دارند.
28- سلول هاي يوكاريوتي 3 نوع RNA پلي مراز دارند؛ كه با شماره هاي I و II و III نشان داده مي شوند.
29- RNA پلي مرازI رونويسي از ژن هاي مربوط به rRNA ها را انجام مي دهد.
30- RNA پلي مرازII رونويسي از ژن هاي مربوط به mRNA و نيز برخي RNA هاي كوچك را انجام مي دهد.
31- RNA پلي مراز III رونويسي از ژن هاي مربوط به tRNA و برخي RNA هاي كوچك ديگر را انجام مي دهد.
32- رونويسي را در طي سه مرحله مي توان نشان داد:
a. در مرحله اول RNA پلي مراز به راه انداز متصل مي شود ( راه انداز قسمتي از DNA است كه به RNA پلي مراز امكان مي دهد كه رونويسي از محل صحيح را آغاز كند)
b. در مرحلهي دوم RNA پلي مراز دو رشتهي DNA را از هم باز مي كند.
c. در مرحلهي سوم RNA پلي مراز همچون قطاري در طول DNA حركت و در مقابل هر نوكلئوتيد آن ، نوكلئوتيد مكمل RNAيي آن را قرار مي دهد تا به جايگاه پايان رونويسي برسد ( جايگاه پايان رونويسي بخشي از DNA است كه بعد از رونويسي از آن ، RNA پلي مراز و DNA و RNA ساخته شده از هم جدا مي شوند و رونويسي پايان مي پذيرد.)
33- در همانند سازي DNA هر از دو رشته به عنوان الگو استفاده مي شود ولي در رونويسي فقط از يكي از دو رشته ي مولكول DNA ( ژن مورد نظر) به عنوان الگو استفاده مي شود.
34- هيچ گاه در يك ژن به طور همزمان از دو رشته در يك قسمت رونويسي صورت نمي گيرد.
35- در بخشي از مولكول DNA ممكن است كه به طور همزمان چندين مولكول RNA رونويسي شوند.
36- نيرنبرگ و همكارانش اولين گروهي بودند كه با استفاده از mRNA رمز DNA را كشف كردند.
37- نيرنبرگ و همكارانش مولكول mRNAي مصنوعي ساختند كه فقط از توالي نوكلئوتيدي يوراسيل تشكيل شده بود ، سپس اين mRNA را در لولهي آزمايشي قرار دادند كه در آن انواع اسيد هاي آمينه و عصارهي استخراجي سيتوپلاسم ( آنزيم هاي ترجمه) وجود داشت؛ در پايان پلي پپتيد ساخته شده را استخراج و تجزيه كردند، اين پلي پپتيد فقط از اسيد آمينهي فنيل آلانين تشكيل شده بود.بنابر اين UUU = يك اسيد آمينه فنيل آلانين اضافه كن مي باشد.(رمز فنيل آلانين بر روي DNA توالي AAA مي باشد.)
38- در كل هر اسيد آمينه حداقل يك رمز روي RNA (يا DNA )دارد.(بسياري از اسيدهاي آمينه بيش از يك رمز دارند)
39- سه رمز پايان وجود دارد كه عبارتند از UAA و UAG و UGA .
40- رمز آغازين هميشه AUG است كه مربوط به اسيدآمينه متيونين است.
41- در ترجمه توالي نوكلئوتيدي mRNA به توالي آمينواسيدي در رشته ي پلي پپتيدي ترجمه مي شود.
42- در ترجمه در اصل زبان نوكلئيك اسيدي به زبان آمينواسيدي ترجمه مي شود.
43- پروتئين سازي در ريبوزوم ها صورت مي گيرد( rRNA ) و tRNA اسيد هاي آمينه را به ريبوزوم حمل مي كند و mRNA اطلاعات را به ريبوزوم حمل مي كند. ( در پروتئين ستزي هر سه نوع RNA شركت دارند)
44- tRNA ساختار برگ شبدري دارد ولي در سلول ساختار اصلي آن شكل L دارد.
45- ضد رمز در برگ مياني قرار دارد و آنتي كدون ناميده مي شود.
46- براي هر اسيد آمينه حداقل يك نوع tRNA وجود دارد.
47- قسمت انتهايي و بدون برگ tRNA جايگاه اتصال آمينواسيد است و توالي انتهايي آن هميشه CCA مي باشد.
48- tRNA ي داراي آنتي كدون GAA ناقل اسيد آمينهي لوسين است و مكمل آن رمز CUU مي باشد.
49- tRNA ي داراي آنتي كدون UAC ناقل اسيد آمينهي متيونين است و مكمل رمز آن AUG مي باشد.
50- tRNA ها ي داراي آنتي كدون هاي ACA و ACG ناقل اسيدآمينهي سيستئين هستند و مكمل رمز آنها UGU و UGC مي باشند.
51- ترجمه داراي سه مرحلهي آغاز ، ادامه و پايان است.
52- فرآيندهاي همانند سازي ، رونويسي و ترجمه هر سه از نوع ساختن و نيازمند به انرژي هستند.
53- ريبوزوم داراي دو زير واحد بزرگ و كوچك است كه زير واحد كوچك خود داراي دو جايگاه به نام هاي A براي آمينواسيد و P براي پلي پپتيد در حال ساخت است.
54- در مرحله ي آغاز ترجمه tRNAي آغازگر (ناقل متيونين) در جايگاه P ريبوزوم قرار مي گيرد و با كدون آغاز ( AUG) رابطهي مكملي برقرار مي كند.سپس زير واحد بزرگ ريبوزوم به زير واحد كوچك متصل مي شود.
55- در مرحله ي ادامه tRNA ي بعدي در جايگاه A قرار مي گيرد و دومين اسيد آمينه با اولين اسيد آمينه پيوند پپتيدي برقرار مي كند، سپس tRNA ي موجود در جايگاه P آزاد شده و جايگاه P خالي مي شود و tRNA ي جايگاه A به همراه رشته ي پلي پپتيدي به اندازه ي يك كدون جابجا شده و جايگاه P را اشغال مي كندو جايگاه A خالي مي شود و tRNAي سوم در جايگاه A قرار ميگيرد و ...
56- در مرحله ي پايان با قرار گرفتن يكي از كدون هاي پايان در جايگاه A ريبوزوم ، زير واحد هاي ريبوزوم و mRNA و رشته ي پلي پپتيدي ساخته شده از هم جدا مي شوند و ترجمه به اتمام مي رسد.
57- يك mRNA مي تواند به طور همزمان توسط چند ريبوزوم ترجمه شود.
58- هر mRNA عمر مشخصي دارد و بعد از مدتي از بين خواهد رفت.
59- در يوكاريوت ها mRNA ي تازه ساخته شده دچار تغييراتي مي شود تا به mRNAي بالغ تبديل شود.
60- يكي از تغييرات mRNA براي بالغ شدن ، كوتاه شدن آن است.
61- اگزون بخشي از DNA است كه رونوشت آن در mRNA باقي مانده و ترجمه مي شود.
62- اينترون بخشي از DNA است كه رونوشت آن در mRNA حذف مي شود و ترجمه نمي شود.
63- اگزون و اينترون مخصوص سلول هاي يوكاريوتي هستند.
64- ژن هاي يوكاريوتي به خاطر داشتن اگزون و اينترون ، گسسته هستند.
65- بيان شدن يك ژن يعني اينكه آن ژن مورد استفاده قرار گرفته است و در اصطلاح روشن است .
66- وقتي ژني مورد استفاده قرار نگيرد ، خاموش است .
67- اينكه در يك زمان مشخص ، كدام ژن روشن و كدام ژن خاموش باشد، تنظيم بيان ژن ناميده مي شود.
68- تنظيم بيان ژن براي پاسخ به شرايط محيطي و نمو مهم است.
69- مادهي ژنتيك همهي سلول هاي بدن ما يكسان است ؛ تفاوت سلول ها به خاطر اين است كه در هر نوع سلول فقط بعضي از ژن ها بيان مي شوند و روشن هستند و بقيهي ژن ها خاموش هستند.
70- آنچه كه فنوتيپ را تعيين مي كند ، نوع پروتئين ها است كه خود محصول ژن هاي روشن هستند.
71- تنظيم بيان ژن ممكن است در سطوح مختلفي مثل قبل از رونويسي يا در هنگام رونويسي و يا بعد از رونويسي صورت بگيرد .
72- معمولا تنظيم بيان ژن در هنگام رونويسي صورت مي گيرد ، چون در اين حالت انرژي كمتري صرف مي شود.
73- تنظيم بيان ژن در هنگام رونويسي يعني اينكه اگر به محصول آن ژن نيازي نباشد از آن ژن رونويسي صورت نمي گيرد .
74- اپران بخشي از DNA است كه شامل دو بخش تنظيم كننده و ساختاري است .
75- بخش تنظيم كننده خود شامل دو قسمت راه انداز و اپراتور است .
76- بخش ساختاري بخشي است كه از روي آن mRNA رونويسي مي شود.
77- بخش ساختاري خود مي تواند شامل چندين ژن باشد .
78- اپران مربوط به سلول هاي پروكاريوتي است و توسط ژاكوب و مونو براي توضيح نحوه ي بيان هماهنگ ژن ها در باكتري ها پيشنهاد شده است .
79- بخش تنظيم كننده ، بيان همزمان ژن ها را كنترل مي كند.
80- اپراتور و راه انداز كنار همديگر هستند .
81- بخشي به نام ژن تنظيم كننده كه ممكن است فاصله ي زيادي از راه انداز داشته باشد باعث توليد پروتئيني به نام مهار كننده مي شود ( پروتئين تنظيم كننده )
82- چسبيدن مهار كننده به به اپراتور ، سدي در جلو RNAپلي مراز ايجاد مي كند كه به راه انداز چسبيده است و بنابر اين ژن خاموش مي شود.
83- در اپران لك با ورود لاكتوز به محيط ، لاكتوز در باكتري به الولاكتوز (عامل تنظيم كننده) تبديل مي شود كه الولاكتوز به مهار كننده (پروتئين تنظيم كننده ) مي چسبد و از اتصال آن به اپراتور جلوگيري مي كند و بنابر اين اپران لك روشن مي شود.
84- در سلول هاي يوكاريوتي ساختار اپران وجود ندارد .
85- در سلول هاي يوكاريوتي فرصت بيشتري براي تنظيم بيان ژن وجود دارد چون DNA در هسته است و از سيتوپلاسم (محل ترجمه) جدا مي باشد.
86- در سلول هاي يوكاريوتي تنظيم بيان ژن ممكن است در سطوح رونويسي يا حتي ترجمه ( در سيتوپلاسم ) صورت گيرد .
(قبل از رونويسي – رونويسي – بعد از رونويسي ) كه اينها در هسته است / (قبل از ترجمه – ترجمه – بعد از ترجمه ) كه اين ها در سيتوپلاسم است .
87- غالبا تنظيم بيان ژن در يوكاريوت ها در هنگام شروع رونويسي صورت مي گيرد.
88- در يوكاريوت ها RNAپلي مراز به تنهايي نمي تواد راه انداز را شناسايي كرده و بر روي آن قرار گيرد و براي اين كار نياز به بخش هاي پروتئيني به نام عوامل رونويسي دارد .
89- عوامل رونويسي بسيار متنوع و متعدد هستند و نقش هاي مختلفي را در تنظيم بيان ژن دارند .
90- گروهي از عوامل رونويسي در ابتدا با راه انداز متصل مي شوند و بعد از آن RNAپلي مراز به آنها مي پيوندد .
91- افزاينده بخشي از مولكول DNAاست كه به كمك عوامل رونويسي متصل به آن (فعال كننده) ، عمل رونويسي را تقويت مي كند .
92- افزاينده بر خلاف راه انداز ممكن است هزاران نوكلئوتيد از ژن فاصله داشته باشد .
93- افزاينده و فعال كننده متصل به آن با تشكيل يك حلقه در DNA در كنار RNAپلي مراز و ساير عوامل رونويسي روي راه انداز قرار مي گيرند و به اين ترتيب عوامل رونويسي متصل به راه انداز فعال شده و ژن روشن مي شود.
94- افزاينده و فعال كننده هستند كه عوامل رونويسي متصل به راه انداز را فعال مي كنند.
95- هر گونه تغيير در DNA را جهش مي ناميم .
96- جهشي كه در سلول هاي جنسي رخ دهد ممكن است به زاده ها منتقل شود.
97- جهشي كه در سلول هاي پيكري رخ دهد فقط بدن خود فرد را تحت تأثير قرار مي دهد و به نسل بعد منتقل نخواهد شد.
98- جهش هايي كه يك يا چند نوكلئوتيد ژن را روي يك كروموزوم تغيير مي دهند ، جهش نقطهاي ناميده مي شوند.
99- در كل جهش هاي نقطهاي دو نوع هستند 1) جهش هاي جانشيني كه در آن در جاي يك نوكلئوتيد ، نوكلئوتيدي از نوع ديگر قرار مي گيرد . 2) جهش هاي افزايشي يا كاهشي كه يك يا چند نوكلئوتيد در ژن حذف يا اضافه مي شود.
100- جهش هاي افزايشي يا كاهشي معمولارمز هاي سه حرفي را به هم مي ريزند .
101- جهشي كه باعث اشتباه خوانده شدن حروف سه نوكلئوتيدي شوند ، جهش تغيير چهارچوب ناميده مي شوند زيرا چهارچوب الگوي خواندن در يك يا دو موضع جابجا مي شود.
102- جهش هاي نقطه اي مي توانند باعث شوند كه الف) پروتئين مورد نظر ساخته نشود ب) پروتئيني متفاوت ساخته شود كه عملكرد متفاوتي داشته باشد.
103- گاه جهش جانشيني بي تأثير است چون بعضي از اسيد هاي آمينه بيش از يك رمز دارند .
مثال: GGU و UGC هر دو رمز هاي مربوط به سيستئين هستند بنابراين اگر در جهش جانشيني به جاي C ، U قرار بگيرد مشكلي پيش نمي آيد .